TCA Cycle and the Electron Transport Chain
Learning Objectives
Evaluate the potential metabolic fates of pyruvate and the signals that control these changes.
Assess the importance of the mitochondria in glucose metabolism.
Describe the key regulatory nodes of the TCA cycle.
Understand the concepts of anaplerosis and cataplerosis and how this can affect TCA cycle efficiency. Predict whether a particular pathway is anaplerotic or cataplerotic.
Explain the differences in efficiency between anaerobic glycolysis and the TCA cycle linked to the electron transport chain.
Recall the key functions of the electron carriers NADH, FADH2 and QH2.
Calculate ATP production from GTP, NADH and FADH2 equivalents.
Understand how mitochondria balance nutrient flux with ATP requirements.
Key Vocabulary and Concepts
Anaplerosis
Cataplerosis
Electron Carrier Molecules (including NADH, FADH2 and QH2)
Proton Gradients and the Electromotive Force
Oxidative Stress, Antioxidants, and Reactive Oxygen Species
The Next Steps in Carbohydrate Metabolism Require Mitochondria
While glucose oxidation to the level of Pyruvate generates the equivalent of 7 ATP molecules2 x NADH (5 ATP equivalents at 2.5 ATP/NADH) and 2 x ATP are generated., full oxidation to CO\(_2\) can yield up to 32 molecules of ATPThe specific amount can vary from 30-32 depending on mitochondrial efficiency.. This process requires transport of pyruvate into the mitochondria where Acetyl-CoA is generated and degraded to CO\(_2\), GTP and reduction of electron carrier molecules. These molecules, NADH and FADH2, are passed to the electron transport chain, generating a proton gradient, which powers the mitochondrial ATPaseThis is a little confusingly named, as an "ase" generally can break down a molecule. In this case the mitochondrial ATP synthase generates ATP from ADP.. Mitochondria are also the only way to catabolize fatty acids, and most Amino Acids. As a membrane-enclosed organelle, many key metabolites are impermeable to mitochondria without transporters. These include cytoplasmic NADH generated during glycolysis as well as PyruvateNADPH cannot cross the mitochondrial membrane, so requires shuttle pathways such as the malate–aspartate and glycerol–phosphate shuttles to indirectly share NADPH between compartments. For more detail about these shuttles see McKenna et al. (2006).. Fortunately there are a variety of transporters and shuttle systems that transport key materiel from the cytoplasm into the mitochondria.
Regulation of Mitochondrial Numbers
Before we start with what happens in mitochondria, its worth taking a step back and considering how mitochondrial amounts are regulated. The presence of mitochondria is essential for aerobic metabolism, and muscle fibers that are rich in mitochondria are able to fully oxidize glucose and other fuels. These are known as Type I muscle fibers, or colloquially as slow-twitch muscle fibersThere is another fast-twitch fiber type that contains less mitochondria called IIA fibers. The number and efficiency of mitochondria in muscle is not static, and can be modified by training. For example endurance training dramatically increases the number of mitochondria and the levels of TCA and ETC enzymes in both rodent and human studies (Holloszy 1967, 2011; Gollnick et al. 1972, 1973). Understanding the molecular mechanisms by which this happens is a major research area both in terms of human performance, and in terms of modifying energy expenditure and promoting healthy aging (for more details see Cartee et al. (2016)). Some cells have few or no mitochondria. In this case they are entirely dependent on glycolysis for ATP production.
The Possible Fates of Pyruvate
As we discussed in the unit on glycolysis, pyruvate has several possible fates. If Alanine levels are low and Glutamate levels are high, Pyruvate can be converted to Alanine via ALTAlanine Aminotransferase.. If there is energy demand, PDHPyruvate Dehdyrogenase, discussed in the next section. is activated and pyruvte becomes Acetyl-CoA. If Acetyl-CoA levels are high, Pyruvate becomes the TCA Cycle intermediate Oxaloacetate via the actions of Pyruvate Carboxylase. If none of these enzymes are activated, Pyruvate is converted by Lactate Dehydrogenase and released from the cell as Lactate. The reversible Lactate Dehydrogenase reaction is:
\[\begin{equation} \label{eq:ldh} Pyruvate + NADH \rightleftharpoons Lactate + NAD^+ \end{equation}\]
Anaerobic Glycolysis
If oxygen is unavailable to the cell, or there are no mitochondria present and lactate is produced, this is the end of anaerobic glycolysis. The reaction to generate lactate uses up the NADH produced during glycolysis. This means that on net one glucose molecule now generates just two ATP equivalentsWalk yourself through this to convince yourself this is correct.. The lactate molecule that is released was once thought to be primarily converted back to glucose by a pathway called the Cori cycleThis will be explained in more detail when we discuss gluconeogenesis. but recent experiments monitoring the flux of lactate shows that a large fraction of lactate is anaplerotic, meaning it is used to increase the number of molecules in the TCA cycle (Hui et al. 2017; Ferguson et al. 2018)Anaplerosis and its inverse, cataplerosis will be described below, but in general is the removal of TCA cycle intermediates..
| Pyruvate Fate | Conditions | Key Enzyme |
|---|---|---|
| TCA cycle | High PDH Activity | PDH |
| Lactate | Low PDH Activity | PDH |
| Alanine | Low Ala, High Glu | ALT |
| Oxaloacetate | High Acetyl-CoA | PCPCPyruvate Carboxylase |
Regulation of Pyruvate Dehydrogenase
Pyruvate DehydrogenaseThis enzyme uses multiple cofactors, including Vitamin B1-derived TPP, Vitamin B2-derived FAD, vitamin B3-derived NAD and Vitamin B5-derived Coenzyme A. Reduced PDH activity is a major cause of Beriberi and Wernicke-Korsakoff syndrome ( which are deficiencies of Vitamin B1. is a mitochondrial enzyme that catalyzes this irreversible reaction:
\[\begin{equation} Pyruvate + NAD^+ + CoA \rightarrow AcetylCoA + CO_2 + NADH +H^+ \end{equation}\]
The two most important products here are Acetyl-CoA, which will enter the TCA cycle and NADH which will go to Complex I of the Electron Transport Chain. As one of the most important molecular decision steps, it makes sense that this enzyme is tightly regulated. Intracellularly, PDH activity is turned on when energy is low, and turned off when energy is available. The specific regulators are shown in Table ↑ and Figure ↑. The first several of these should be familiar to you, as increased substrate, and decreased energy (as sensed by low ATP, high ADP and low NADH, Acetyl-CoA) all promote PDH activityNow you may be thinking, I thought the key determinant of aerobic respiration was oxygen availability! It may not be clear yet, but once you have read these notes, try to come back to this and think about how low oxygen levels would result in changes in PDH activity. Ca2+ is an activator of PDH activity, and as we will describe, also plays a role in activating several other TCA Cycle enzymes. This is because, when muscles contract they release Calcium. This allows for mitochondria to respond to contraction by generating more ATP.
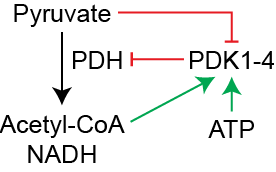
Much like PFK2 and Pyruvate Kinase, PDH is Inhibited by Protein Phosphorylation. The phosphorylation of PDH is regulated by a specific protein kinase (PDH Kinase or PDK) which adds a phosphate group to PDH and reduces its activity (see Figure ↑). The overall mechanism of regulation of PDH therefore is a combination of direct inhibitors of PDH (NADH and Acetyl-CoA), activators of the inhibitory kinase (ATP, Acetyl-CoA, NADH) and activators of the activating phosphatase (Ca2+). Unlike PFK2 and Pyruvate Kinase, PDK activity is regulated mostly at the metabolite level and not acutely by hormonal signals and protein phosphorylation.
Pyruvate Dehydrogenase Kinase is regulated transcriptionally. While the activity of PDK is primarily regulated by metabolites, the number of PDK enzymes can be induced by several signals. Here are a couple, as you read these take a moment to think why this stimuli would alter PDK levels, and what would be the effects on glycolysis, gluconeogenesis and mitochondrial respiration. Remember more PDK means less PDH activity:
Glucocorticoids induce PDK4 transcription via FOXO (Connaughton et al. 2010).
Starvation induces PDK4 (Wu et al. 1998).
Insulin reduces PDK4 levels, insulin resistance elevates PDK4 levels (Harris et al. 2001).
Hypoxia (low oxygen) induces PDK1 via HIF1\(\alpha\) (Kim et al. 2006).
| Regulator | Effect | Mechanism |
|---|---|---|
| Pyruvate | Positive | Inactivates PDK |
| ADP | Positive | Inactivates PDK |
| ATP | Negative | Activates PDK |
| Acetyl-CoA | Negative | Activates PDK, Inhibits PDH |
| NADH | Negative | Inhibits PDH |
| Ca2+ | Positive | Activates PDH Phosphatase |
Acetyl-CoA also enters the TCA Cycle after \(\beta\)-oxidation. While we have focused so far on carbohydrate metabolism, now is a good time to talk briefly about lipid oxidation. Unlike carbohydrates, when fatty acids are broken down they produce Acetyl-CoA, not PyruvateThey also generate equal amounts of NADH, FADH2 which we will discuss later. This means that in humans, fatty acids enter the TCA cycle here. If fatty acids are oxidized in the liver, but the TCA Cycle is not activated, those extra Acetyl-CoA molecules are released as ketone bodies, which can be used by other tissues, after re-conversion back into Acetyl-CoA.
Some amino acids are also converted into Acetyl-CoA. As we will learn in the gluconeogenesis and amino acid catabolism lectures, some amino acids, depending on the circumstances are able to become converted into glucoseThese are the glucogenic amino acids.. Others are converted into Acetyl-CoA and are known as the ketogenic amino acids. These amino acids can only be oxidized into energy by entering the TCA cycle as Acetyl-CoA, but cannot become glucoseLater we will learn that the exclusively ketogenic amino acids are Leucine and Lysine. Phenylalanine, Isoleucine, Threonine, Tryptophan and Tyrosine are partially glucogenic and partially ketogenic..
The TCA Cycle Products
The TCA Cycle/ETC completely oxidizes Acetyl-CoA to CO2. One cycle, using one molecule of Acetyl-CoA generates the following:
| Product | \(\rightarrow\) | ATP |
|---|---|---|
| 3 NADH | \(\rightarrow\) | 7.5 ATP |
| 1 FADH2 | \(\rightarrow\) | 1.5 ATP |
| 1 GTP | \(\rightarrow\) | 1 ATP |
| Total | 10 ATP |
if we include the NADH generated by Pyruvate Dehydrogenase, that means that Pyruvate oxidation results in 12.5 molecules of ATP.@ Based on what we have discussed in this unit and in the glycolysis unit, try to determine how much ATP is generated from one molecule of glucoseFor a slightly bigger challenge, consider that palmitate oxidation generates 8 Acetyl-CoA, 7 NADH and 7 FADH2 molecules, but requires 2 ATP molecules for activation. Consider the energy yield from a C16:0 (Palmitate) fatty acid, compared to glucose.. Compare this to the two molecules of ATP generated by anaerobic glycolysis and it should make sense why we breathe heavier when we exercise.
NADH and FADH2 Are Substrates for the Electron Transport Chain
Once Acetyl-CoA is generated by PDH, or by the breadown of amino or fatty acids, the TCA cycle functions to extract electrons from these substrates and pass them along to the electron transport chain. This is done by transferring electrons from TCA intermediates to electron carrier molecules including NADH and FADH2. These molecules are “charged” by these electrons, and then transfer them to Complex I (for NADH) or Complex II (for FADH2). The electron transport chain takes these charged carriers, and transfers the energy through a series of complexes. After Complex I or II, electrons are next transported to another carrier called Ubiquinone (or Coenzyme Q) which takes them to Complex III.@ Between complex III and IV is a final carrier called Cytochrome c. As electrons are passed between electron carriers, complexes I, III and IV result in proton ions being pumped out of the mitochondrial cristae, resulting in a gradient of protons. The final step at complex IV reduces oxygen into water. Oxygen is the final electron carrier and is essential for the complete oxidation of the electron carrier molecules.
| Molecule | () | ATP |
|---|---|---|
| 1 NADH | () | 2.5 ATP |
| 1 FADH2 | () | 1.5 ATP |
| 1 GTP | () | 1 ATP |
The Electron Transport Chain is Coupled to ATP Production
As a result of the reactions in Complex I, III and IV, protons are pumped from the inside of the mitochondria to the inner-membrane spaceMitochondria have two membranes an inner membrane and an outer membrane.. This generates a proton gradient with more protons on the outside of the inner membrane than on the inside. This gradient drives a proton-coupled pump called ATP Synthase. If the proton gradient is established, and sufficient ADP levels are present inside the mitochondria, ATP Synthase catalyzes the production of ATP from ADP.@
The three electron carrier molecules we have described are FAD, NAD and CoQCytochrome c is a protein not a small molecule.. These all need to be available in the mitochondria to allow electrons to flow from the TCA cyle through the ETC.@ These molecules are reduced by the actions of the TCA cycle then oxidized back to their original form by the ETC.@ All three of these can be generated endogenously, but NAD and FAD are usually generated from vitamins (see Table ↑). It has been suggested that as we age, we are less able to generate NAD, and several preclinical trials are underway to test whether NAD supplementation may slow aging in humans (reviewed in Rajman et al. (2018)) .Coenzyme Q is generated endogenously using some enzymes of the steroid biosynthesis pathway, and inhibitors of this pathway (such as statins) have been suggested to result in CoQ deficiency (reviewed in Quinzii et al. (2007)).
| Carrier | Source |
|---|---|
| NAD | Vitamin B3 |
| FAD | Vitamin B2 |
| CoQ | Not Considered a Vitamin |
The TCA cycle is both anabolic and catabolic. While the TCA cycle is primarily thought of as a catabolic pathway, it is also a source of many biosynthetic precursors. For example, Citrate can be exported from the mitochondria and converted into Acetyl-CoA for fatty acid synthesis. Other TCA cycle intermediates are used to generate amino acids, heme and nucleotides. We will discuss this in more detail in the section on anaplerosis and cataplerosis below.
Regulation of the TCA Cycle
The TCA/ETC cycle has to be tightly controlled. This happens in three levels. The first is that the entire system is driven by ATP demand. Think of this aspect of metabolism more as being pulled by the need, rather than being pushed by substrates. The second level of regulation is how many TCA cycle intermediates are present. Since this is a cycle, where the starting substrate (oxaloacetate) is regenerated, having more oxaloacetate molecules in the cycle will increase efficiency. The third level of regulation is regulation of the TCA cycle enzymes themselves. This can be allosteric (increased activity with low ATP/NADH), transcriptional or post-translational.
The ETC is Driven by ATP Demand
Many metabolic pathways are driven by the nutrients that flow in. An example of this is that glycolysis occurs more rapidly when more glucose is present. This is facilitated by feed-forward mechanisms where more glucose results in higher activity of PFK (via F26bP) and Pyruvate Kinase (via F16bP). The electron transport chain, however, is primarily regulated by demand, rather than supply. In this case, demand means high levels of ADPbecause of breakdown of ATP to ADP.. In the absence of ADP, electrons from NADH/FADH\(_2\) do not get transported to the final electron acceptor (O\(_2\)). If ADP levels are high, that indicates that ATP needs to be synthesized and the electron transport chain is active.
Changes in TCA Cycle Intermediates
The TCA cycle regenerates Oxaloacetate. Unlike glycolysis, which starts with glucose and ends with Pyruvate, the TCA cycle takes in Acetyl-CoA and after one round of the cycle, is left with Oxaloacetate. That means that other than Acetyl-CoA, the cycle is self-replenishingOne analogy for this is that the TCA Cycle is like a subway system, and Oxaloacetate is like a subway car. You need it to get from point A to point B, but you dont use up the car.. As you might suspect, having more Oxaloacetate can mean there is more efficient Acetyl-CoA metabolism. While normal TCA cycle function as we have been describing does not alter these levels, there are several important processes that can affect this. One example is gluconeogenesis, which extracts Oxaloacetate (via the activity of an enzyme called PEPCK) to form glucose. Another example is that biosynthesis of some amino acids uses up TCA cycle intermediates. The process by which TCA Cycle intermediates are removed is known as cataplerosis. This process is extremely important for growth as it is a source of many building blocks for cells (for more details see Inigo et al. (2021)).
The opposite process, in which TCA cycle intermediates are generated is known as anaplerosis. These can derive from Pyruvate, or from the breakdown of amino acidsAmino Acid catabolism will be covered later in the course, so here we will focus on anaplerosis from Pyruvate.. The most important enzyme here is called Pyruvate Carboxylase. This enzyme performs the following irreversible, ATP consuming reaction:
\[\begin{equation} \label{eq:pcx} Pyruvate + Bicarbonate + ATP \rightarrow Oxaloacetate + ADP + Pi\nomenclature{Pi}{Inorganic Phosphate} \end{equation}\]
There are two important roles of Pyruvate Carboxylase, one of which is to increase TCA Cycle intermediates. The second is to generate Oxaloacetate for gluconeogenesisWe will discuss this in a couple of lectures.. The activity of Pyruvate Carboxylase is positively regulated by Acetyl-CoA. Since Acetyl-CoA is not directly anaplerotic, this mechanism balances flow of “passengers” (Acetyl-CoA) to the number of “trains” (Oxaloacetate). Extending the metaphor, if there are too many passengers, Pyruvate Carboxylase results in more trains being put into service. As we will discuss later in the section on gluconeogenesis, Acetyl-CoA (the major metabolite of fatty acid oxidation) is unable to become glucose, meaning that while fatty acids can promote gluconeogenesis, the process is still reliant on other precursors such as lactate, alanine and glycerol.
Allosteric Regulation of the TCA Cycle
As we discussed above, the ETC is inactive unless there is ATP demand. It is therefore imperative that the ETC can feed back to the TCA cycle and stop NADH/FADH\(_2\) production if these activated electron carriers are not needed. The primary mechanism by which this occurs is negative allosteric regulation described in Table ↑. The signals of high energy availability are ATP, NADH and FADH\(_2\).
| Enzyme (step) | Activators | Inhibitors |
|---|---|---|
| Citrate synthase (1) | OAA, AMP | NADH, FADH\(_2\), ATP |
| Isocitrate dehydrogenase (3) | ADP, Ca\(^{2+}\) | NADH, ATP |
| \(\alpha\)-ketoglutarate dehydrogenase (4) | Ca\(^{2+}\) | NADH |
You should note from Table ↑ that these enzymes are broadly activated by two things, low energy levels (AMP or ADP) and Calcium increases. Calcium plays a key role here, because in skeletal muscle Calcium release causes muscle contraction. By activating the TCA cycle, Calcium couples the initiation of muscle contraction to the replenishment of ATP levels. By the same token, the TCA cycle is inhibited when there is a buildup of high energy molecues (NADH, FADH\(_2\), ATP) either when there is a lack of oxygen or sufficient energy stores.A high NADH/NAD+ ratio, such as that as seen in hypoxia or ischemia, inhibits isocitrate dehydrogenase and \(\alpha\)-ketoglutarate dehydrogenase. This slows the TCA cycle, reducing NADH generation when the electron transport chain is backed up. The result is accumulation of upstream intermediates and diversion toward anaerobic metabolism (lactate production). In severe ischemia, for example after an ischemic heart attack, this contributes to cytosolic acidification and impaired ATP synthesis.
Anemia and the TCA Cycle
Several enzymes in the TCA cycle as well as the electron transport chain require iron as a cofactor (e.g. Aconitase and Complex II). In addition to its role as a cofactor, iron is essential for the delivery of oxygen to cells, which is mediated by iron-containing hemoglobin, and for oxygen transport within muscle cells, which is mediated by iron-containing myoglobin. For these reasons, low iron levels can impair TCA cycle function and aerobic metabolism. This is one of the reasons why anemia can cause fatigue and exercise intolerance. Anemia can also be caused by other chronic diseases such as chronic kidney disease, inflammatory bowel disease and cancer, and this can contribute to fatigue in these conditions. In the United States anemia affects 9.3% of the population, and is 2.4x more common in women than men (Williams et al. 2024). Globally anemia is the most common nutritional deficiency, affecting 1.62 billion people or about a quarter of people worldwide (World Health Organization 2025).
Oxidative Damage Can Result from Superoxide Production
The terminal electron acceptor of the TCA cycle and electron transport chain is oxygen. This is the reason why aerobic metabolism requires a supply of oxygen. The oxygen is converted to water by Complex IV of the ETC.@
\[\begin{equation} \label{eq:civ} 4 CytC_{red} + 4H^+ + O_2 \rightarrow 4 CytC_{ox} + 2 H_2O \end{equation}\]
While this process is efficient most of the time, due to the high reactivity of oxygen, approximately 2-4% of the time, rather than being reduced to water, oxygen can be partially reduced into a superoxide or peroxide molecule. These reactive oxygen species such as \(HO_2^-\) can cause cellular damage by reacting with proteins and lipids in the cell. This process can also be accelerated when NADH or FADH2 build up. To defend against this, mitochondria express an enzyme known as superoxide dismutase (SOD) which scavenges these superoxides and converts them to hydrogen peroxide and then via an enzyme known as catalase back into water. These reactions are described in reactions ↑ and ↑ below:
\[\begin{equation} \label{eq:sod} 2 HO_2^- \rightarrow O_2 + H_2O_2 \end{equation}\]
\[\begin{equation} \label{eq:catalase} 2 H_2O_2 \rightarrow 2 H_2O + O_2 \end{equation}\]
Overwhelming this system has been linked to a wide variety of chronic diseases including Alzheimer’s, Parkinson’s disease, Cancer and Diabetes. One of the benefits of chronic physical activity is to increase the expression of SOD and catalaseFor those interested in micronutrients, catalase is an iron-dependent enzyme and SOD is either a mangangese or copper dependent enzyme, and deficiencies in any of these minerals can enhance oxidative damage.. By the same token, antioxidant vitamins such as Vitamins C and E can also reduce the potential damage of reactive oxygen species generated by the mitochondria. For more information about reactive oxygen species see Turrens (2003).
Reflection Questions
A marathon runner sustaining aerobic exercise and a sprinter working anaerobically both start with glucose. Trace the fate of pyruvate in each scenario, explaining how PDH activity, PDK regulation, and mitochondrial ATP demand differ between these two contexts. Include the role of calcium signaling in your answer.
During prolonged fasting, the body relies heavily on fatty acid oxidation, which generates large amounts of Acetyl-CoA but cannot contribute carbons to glucose. Using your understanding of anaplerosis, cataplerosis, and pyruvate carboxylase regulation, explain how gluconeogenesis can still be sustained during fasting, and describe the specific role Acetyl-CoA plays in coordinating this process.
A patient with iron-deficiency anemia reports severe fatigue during exercise. Their physician recommends high-dose antioxidant supplementation (vitamins C and E) to reduce oxidative stress. Analyze the multiple steps in aerobic metabolism where iron deficiency impairs function, and evaluate whether antioxidant supplementation would be expected to address the primary mechanism of fatigue in this patient.