Metabolic Control Systems
Learning Objectives
Explain what a rate limiting enzyme is, what a committed enzyme step is and what a reversible reaction is.
Predict the differences in speed and persistence of allosteric, post-translational and transcriptional regulation of metabolism.
Describe the role of cellular transport in macromolecular regulation. Understand the differences between active and passive transport.
Key Terms and Concepts
Activation Energy
Allosteric Regulation
Cofactor
Compartmentalization
Concentration Gradient
Enzyme
Feedback Inhibition
Isoenzymes
Post-Translational Modification
Rate Limiting Enzyme
Transcription Factor
Transporters (Active and Passive)
Control of Metabolic Flux
Cells need to control the rates at which nutrients are taken up, stored, or used and there are several ways by which this occurs. Here we will review the biochemistry of both nutrient transport and enzyme function. Understanding these concepts will be very important to understanding how the metabolic pathways we will discuss are controlled.
Cellular Transport Systems
First we will describe the ways in which cells control nutrient permeability. Most of the nutrients we will discussThe exceptions are sterols and some other lipids are unable to pass through the plasma membrane of the cell. Allowing or denying access to a nutrient is one way by which cells can control nutrient metabolism. Without these transport mechanisms we would be unable to absorb digested food, or transport nutrients from cell to cell. While we normally think of transporters as getting nutrients into or out of a cell, they are also important within cells, for example getting pyruvate into the mitochondria, or storing calcium in the endoplasmic reticulum.
Types of Membrane Transporters
Membrane transporters are generally fairly specific for the molecule they transport. For example GLUT4 transports glucose, but GLUT5 transports fructose. Transporters can broadly be separated into two major types, passive transporters and active transporters. These can be differentiated by considering whether they work with or against the concentration gradient, with active transporters typically working against the concentration gradient.
Passive transporters allow for nutrients to pass down a concentration gradient into the cell. As an example, the liver expresses a glucose transporter named GLUT2. Glucose can either enter the liver (if there is more glucose in the blood than the liver) or exit the liver (if the reverse is true). Passive transporters will only allow a nutrient to enter a cell if there is less of the nutrient in a cell than in the blood. This is quite efficient for disposing of excess nutrients, such as after a meal, but is not effective in storing things away against a concentration gradient. It may seem like passive transporters are not regulated, but as we will see in the case of GLUT4, the amount of transporters at the cell surface can be controlled by cell signalingif you want to jump ahead, here is a review on that process (Leto and Saltiel 2012). The rate of a passive transport is defined by three things, the gradient of the transported molecule, the number of transporters at the relevant membrane, and the efficiency of the transporter.
Active transporters can force nutrients into a cell against the concentration gradient. These transporters function like pumps and have to use energy of one sort or another to force the molecule into the cell. You may think that this is a bad idea, but there are lots of examples where this matters physiologically. One example is retaining salt. If your kidneys weren’t actively retrieving sodim out of urine and back into the blood, then you would rapidly lose osmotic pressure in your blood. The key is to think about the concentration inside or outside the cell, and if you are pushing against the transport gradient, you need active transport.
Powering Active Transport
Active transport requires energy of some type. This energy can come from several sources such as ATP, other concentration gradients, or even light. Some examples are described in Figure ↑. The key to controlling the rate of these transporters is not only the concentration gradient of the transported molecule, but also the levels (or gradient) of the powering force. In the cases where molecules are co-transported they can either be pulled in simultaneously (this is known as a symporter) with the molecule of interest as shown on the left of Figure ↑, or can be exchanged where one molecule exits, powering the entry of the molecule of interest (this is known as an antiporter). A classic example of a symporter is the sodium:glucose cotransporter SGLT1, which brings sodium into the cell down its concentration gradient (a gradient maintained by the basolateral sodium:potassium ATPase) to simultaneously power the uptake of glucose from the gut lumen, even against glucose’s own concentration gradient. This allows for efficient carbohydrate uptake in a mealSGLT2 does a similar thing, retrieving glucose from urine back into the blood. Therefore inhibiting SGLT2 prevents glucose retrieval back to the blood, and is the target of several drugs which try to lower blood glucose in diabetics. The trade names for these drugs include Invokana, Farxiga and Jardiance. even if the gastrointestinal cells have similar or higher glucose levels to the gut lumen.
Transporters themselves can be regulated. This is often done by changing the number of transporters at the cell surface, or by changing the activity of the transporter. For example, in the case of GLUT4, insulin stimulates the translocation of GLUT4 from intracellular vesicles to the cell surface, increasing the amount of glucose that can be taken up by the cell. This is a common mechanism by which cells can rapidly respond to changes in nutrient availability. Other transporters can be regulated by changing their activity, for example by phosphorylation. See the section below Integrated Control of Metabolism by Regulation of Enzymes for more details on how this happens.
Enzymes
Thermodynamics
Every chemical in the body has a certain amount of energy. When we eat, some of this chemical energy is converted into ATP to allow for function. This is known as catabolism. When we are storing nutrients, we use ATP to generate higher energy molecules such as fats or glycogen. This is known as anabolism. Every molecule in our body has a set amount of energy and a chemical reaction can be considered endothermic (requiring energy) or exothermic (releasing energy), depending on whether the reactants or products have higher energy. The levels of these metabolites at equilibrium can be calculated with the following equation where K\(_{eq}\) is the Eqiulibrium constantThe square brackets mean the concentration of A or B:
\[\begin{equation} K_{eq}=\tfrac{[B]}{[A]} \end{equation}\]
The equilibrium constant can be calculated from the free energy of the reactants and products.
\[\begin{equation} \Delta G^{'}_{o} = - R T \ln K_{eq} \end{equation}\] \[\begin{equation} \Delta G^{'}_{o} = G^{'}_{o} (products) - G^{'}_{o} (reactants) \end{equation}\]
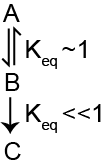
Some reactions have products with very similar energy levels and the balance between the reactants and the products is based primarily on their concentrations. This is known as an equilibrium reaction which would have a K\(_{eq}\) of near to 1. If a reaction releases a large amount of energy, it is often an irreversible or committed stepThis would have a large, negative \(\Delta G\). Because so much energy is released in the forward direction, the reverse reaction is effectively impossible under cellular conditions, so once this reaction happens, there is no going back. If you think about the metabolic pathway in Figure ↑, this would mean that once you proceed through step 2 to make C you cannot go back to B. Given the free energy (\(G^{'}_{o}\)) and concentration of the reactants and products in a reaction you can calculate the \(\Delta G\) and equillibrium constant for a reaction and estimate whether it is reversibleThere is a good blog post explaining how the steady state \(\Delta G\) is determined on this basis at http://sandwalk.blogspot.com/2007/10/aldolase-reaction-and-steady-state.html under normal conditions.
Enzyme Kinetics
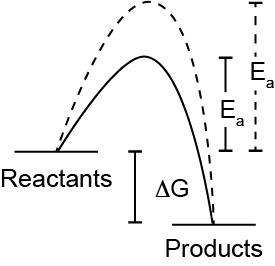
Without enzymes, many reactions occur very slowly due to the activation energy needed for the reaction to occur. Enzymes increase the rate of a chemical reaction by reducing the activation energy required for a reaction to occur. This does not change the equilibrium constant, it just allows the reaction to reach equilibrium faster. This is sketched out in Figure ↑, note that \(\Delta G\) is not changed, but the dashed line has a higher activation energy, and therefore slower reaction rate than the solid line.
Most metabolic pathways are controlled by altering the rates at which metabolites are converted to final products. The overall rate of a metabolic pathway is controlled by the rate-limiting stepthe rate-limiting step is generally the slowest step of a pathway. As an example, in glycolysis the rate-limiting step is catalysed by phosphofructokinase-1. In a linear pathway, the speed of this step’s enzymatic reaction controls the overall rate. Quite often the rate-limiting enzyme is an important point of regulation, as adjusting the speed of this reaction can speed up or slow down and entire pathway.
Reaction rates increase in rate as the concentration of substrates increase until the enzymes are saturated (see the solid line in Figure ↑). This is known as Michaelis-Menten kinetics. The reaction rate constant (k) and rate can be calculated from the activation energy with these equationsA and R are constants, T is temperature and e is Euler’s number. \(K_{m}\) is the Michaelis constant for an enzyme.:
\[\begin{equation} k = A e^{-\frac{E_{a}}{RT}} \end{equation}\] \[\begin{equation} rate = k\tfrac{[Reactant][Enzyme]}{[Reactant] + K_{m}} \end{equation}\]
If products build up the reaction becomes more complex and now looks like this where K\(_{p}\) is the binding constant for the product:
\[\begin{equation} rate = k\tfrac{[Reactant][Enzyme]}{[Reactant] +K_{m}\left \{ 1 + \frac{[Product]}{Kp} \right \}} \end{equation}\]
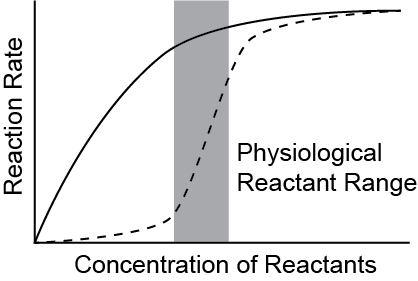
Allosteric regulation is another way by which enzymes can control reaction rates. Allosteric enzymes are generally multi-subunit enzymes that change their K\(_{m}\) as more substrate binds. An example of this is the dashed line in Figure ↑. This has several advantages in terms of regulation. One advantage is that the reaction rate can be effectively zero or at maximum in a much narrower range, bracketing the actual range of substrates present physiologically. Another advantage is that allosteric activators or inhibitors can shift the curve to the left or right, to effectively increase or decrease the reaction rate. This is a common mechanism by which the activity of rate-limiting enzymes are regulated.
On the basis of these equations, reaction rates (and the rate of a particular metabolic pathway) can be increased by several thingsSome examples include: decreasing the activation energy; increasing the amount of the reactants; decreasing the amount of the products; increasing the number of enzymes; decreasing the K\(_m\) of the enzyme; or shifting the substrate sensitivity of the allosteric enzyme.. Try to convince yourselves how this happens based on the equations listed above. Can you think of any other things that would affect pathway flux?
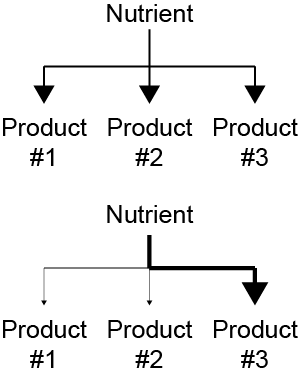
While linear flow through a pathway is important, another aspect of pathway control is how the fate of a particular nutrient is decided. This is illustrated in Figure ↑. In the example on top the nutrient would be equally distributed between three products, but in the bottom example, by adjusting the rates of the specific pathways, a nutrient can be directed to a particular product. At several points during this class, we will describe how the fates of particular metabolites are controlled by the relative rates of metabolic pathways.
Cofactors
Many enzymes require non-protein helper molecules to catalyze their reactions. These are known as cofactors. Table ↑ lists some important cofactors, and the dietary vitamins from which they are derived. Other important roles of vitamins, for example in activating molecules like Co-enzyme A and electron carrier molecules like NADH will be described later in the semester. A lack of a dietary source of a cofactor can often impair the activity of an enzyme.For example, a lack of vitamin B1 (thiamine) can impair the activity of pyruvate dehydrogenase, which is important for converting pyruvate to acetyl-CoA. This can lead to a condition known as Wernicke-Korsakoff syndrome, which is characterized by confusion, ataxia and ophthalmoplegia. This is all because pyruvate dehydrogenase cannot function properly.
| ll@ Cofactor | Source |
|---|---|
| TPPTPPThiamine PyrophosphateVitamins!Vitamin B1 (Thiamine) | Vitamin B1 |
| Pyridoxal PhosphateCofactors!Pyridoxal Phosphate (PLP)Vitamins!Vitamin B6 (Pyridoxine) | Vitamin B6 |
| BiotinVitamins!Vitamin B7 (Biotin) | Vitamin B7 |
| THFTHFTetrahydrofolateVitamins!Vitamin B9 (Folate) | Folic Acid |
| Iron | Dietary Fe |
| Selenium | Dietary Se |
Isoenzymes, Compartmentalization and Tissue Specificity
Enzymes can be tissue specific. This means that the same reaction can be catalyzed by different enzymes in different tissues. These are known as isoenzymes or isozymes. An example of this is lactate dehydrogenase, which has several isoforms that are expressed in different tissues. The liver isoform is more active at lower pH than the muscle isoform, which is more active at higher pH. This means that the liver can continue to produce glucose from lactate even when the blood pH is low, such as during exercise. This is an important concept to understand, as it allows for different tissues to have different metabolic priorities, and will be discussed in more detail later in the course.
Cells are not just a bag of enzymes. They are highly organized structures with different compartments that have different functions. This is known as compartmentalization. For example, the mitochondria are the site of oxidative phosphorylation, while the endoplasmic reticulum is the site of protein synthesis and lipid metabolism. This allows for different metabolic pathways to occur in different parts of the cell, which can help to control the rates of reactions and prevent unwanted side reactions. Compartmentalization also allows for the separation of metabolic pathways that may otherwise interfere with each other, such as glycolysis and gluconeogenesis.Try to think of another example of a metabolic reaction and how it might be compartmentalized to a specific tissue or a subcellular location.
Integrated Control of Metabolism by Regulation of Enzymes
What we do (or do not do) with nutrients is largely governed by the activity of transporters and enzymes. There are several ways in which enzymes are regulated, both based on intracellular and extracellular signals. An example might be that a lack of intracellular ATP causes an increase in ATP producing pathways such as glycolysis. On the other hand, low circulating blood glucose levels may work to stop a glucose consuming process such as glycolysis. We will discuss this in detail throughout the class, but some of the hormones we will discuss in this course that are particularly important are listed in Table ↑:
| Hormone | Main Function |
|---|---|
| Insulin | Reduces blood glucose and lipid levels |
| Adrenaline | Increases blood flow, nutrients to muscle |
| Glucagon | Increases blood glucose levels acutely |
| Cortisol | Increases blood glucose levels chronically |
| GH/IGF-1 | Promotes protein synthesis and bone growth |
| Testosterone | Promotes protein synthesis |
| Leptin, GLP-1 | Suppresses appetite |
| CCK, Gastrin, Secretin | Regulation of digestion |
Hopefully these hormones, how they work and how they are regulated is material you are familiar with from previous classes. If not, or you want a refresher, check out the Endocrine Regulation of Macronutrient Metabolism handout also available on Canvas.
Allosteric Regulation
As noted above, one way by which enzymes are regulated is by allosteric regulation. This is a common mechanism by which the activity of rate-limiting enzymes are controlled. Allosteric enzymes have multiple subunits and can change their conformation when a metabolite binds to them. This can either increase or decrease the activity of the enzyme, depending on the metabolite that binds. Allosteric regulation is often rapid, and can be reversed quickly, so it is a good way to control metabolic pathways in response to changes in substrate or product levels.An example of this is phosphofructokinase-1, which is allosterically activated by AMP and inhibited by ATP. This means that when ATP levels are high, glycolysis is slowed down, but when ATP levels are low, glycolysis is sped up.
Feedback Inhibition
A particularly common and important case of allosteric regulation is feedback inhibition (also called end-product inhibition), in which the final product of a metabolic pathway inhibits an enzyme near the beginning of that same pathway, most often the committed step. This is an efficient design: when the end product is already abundant, the cell shuts the pathway down at its entry point rather than wasting substrate and energy making intermediates it does not need. A good metabolic example is the control of glycolysis by the cell’s energy status. The purpose of glycolysis is ultimately to generate ATP, so when ATP is already plentiful there is no need to run the pathway. Accordingly, ATP allosterically inhibits phosphofructokinase-1, the committed step of glycolysis, slowing glucose breakdown; when ATP falls and AMP rises, this inhibition is relieved and glycolysis speeds back up to replenish ATP. In effect, the pathway’s own product reports back on whether the pathway needs to keep running.
Post-Translational Modification
One common way by which enzymes are regulated is by the modification of existing proteins. One common example is protein phosphorylation. In this example a phosphate molecule is attached to an existing protein, which could increase or decrease its activity. This is often reversible, so a good analogy is that post-translational regulation is like flipping a switch for an enzyme on or off. This can occur fairly rapidly, and is not a permanent change.An example of this is that in response to glucagon or adrenaline, glycogen phosphorylase is phosphorylated, which increases its activity and allows for glycogen breakdown to glucose. This is a good way to rapidly change the activity of a pathway in response to a stimulus, such as low blood glucose levels or exercise.
Transcriptional Regulation
Another way to change the activity of a pathway is to selectively change the number of enzymes. If this is done at the messenger RNA level, it is known as transcriptional regulation. This is because transcription is the process by which new mRNA is made. By increasing or decreasing the rate of mRNA (and eventually protein) production, the cell can respond to a stimulus to make more or less of a particular protein.An example we will discuss in this course is that when stress levels are high, the brain responds by increasing appetite. The regulation of transcription is often controlled by transcription factors, a class of proteins that can bind to selective sites of DNA and recruit the machinery to make (or prevent the making) of mRNA. For a really great review with more details on transcription factors, take a look at Lambert et al. (2018). These kinds of changes are slow, energetically costly and difficult to reverse. They represent a chronic response, and are not appropriate for short term modifications. The relationship between allosteric, post-translational and transcriptional regulation is demonstrated in Figure ↑. Reflect on an example of metabolic regulation that you can think of. Then consider the timescale by which the changes happen, and try to think what would be the most appropriate mechanism to alter metabolism.
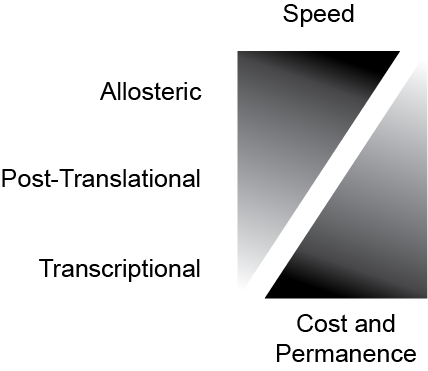
Reflection Questions
The liver takes up glucose from the blood after a meal, when blood glucose is higher than the glucose concentration inside the liver cell. In contrast, the cells lining the small intestine must absorb glucose from the gut lumen even when those cells already contain more glucose than the lumen does. For each situation, decide whether passive or active transport is required, name a transporter from this chapter that could do the job, and explain what powers glucose uptake in the case where it must occur against the concentration gradient.
A drug company develops three candidate compounds targeting the same metabolic enzyme: one is an allosteric inhibitor, one blocks the phosphorylation site used by a kinase, and one suppresses the gene’s transcription factor. A patient needs rapid correction of a dangerously elevated metabolite. Analyze which compound would be most appropriate for acute treatment and which would be better suited for long-term management, using your understanding of the timescales and reversibility of each regulatory mechanism.
Feedback inhibition most commonly acts at the committed step of a pathway rather than the rate-limiting step, yet these are often the same enzyme. Evaluate why targeting the committed step (rather than any slow step) is the more logical control point for a cell. Use a specific example from carbohydrate or lipid metabolism to support your argument.
Fatty acid synthesis occurs in the cytoplasm while fatty acid oxidation occurs in the mitochondria, yet both pathways share several intermediates. Explain how compartmentalization prevents these opposing pathways from running simultaneously in a futile cycle, and identify the transport step that serves as the key regulatory gate between them.