Lipid Digestion and Absorption
Learning Objectives
Identify the roles of Lingual Lipase, Gastric Lipase and Pancreatic Lipase including their locations and specific roles on lipid digestion in the stomach
Understand the role of bile salts in the formation of micelles, and explain how diseases of bile formation can affect an individual
Explain the transport mechanisms of lipids across the apical and basolateral membranes, and how they differ between lipid classes
Describe the re-esterification processes in the enterocyte
Explain the nature, formation and fate of the chylomicron and its role in lipid transportation
Key Vocabulary and Concepts
Micelle
Lipases and Colipase
CCK and Secretin
Gallbladder and Bile Synthesis
Primary, Conjugated and Secondary Bile Salts
Dietary Lipids and Dietary Intake
The three main dietary lipids we ingest are triglycerides, phospholipids and cholesterol. Of these, on average we consume much more in terms of triglycerides (95g for men, 65g/day for women) than phospholipids (1-2g/day) or cholesterol (300 mg/day). For the average person that’s about 1/3 of their total caloric intake (National Center for Health Statistics 2017). Dietary lipids are also our only source of the essential \(\omega\)3 and \(\omega\)6 fatty acids. Furthermore several lipid soluble vitamins are carried and absorbed along with lipids. These include vitamins A, D, E and K, so impairments in lipid absorption can affect their absorption as well.
Lipid Digestion in the Upper Digestive Tract
There is substantial debate about whether lipids can be tasted, in the way that we have specific receptors in our tongue that can sense sweet, bitter, sour, salt, and umami flavors. While there are fatty acid receptors on the tongue, at this stage most lipids are in the triglyceride form. Rather lipid flavor is thought to be a combination of several fatty acid receptors, along with the mechanical sensation of lipids in the oral cavity (DiPatrizio 2014).
Lipid digestion, relative to carbohydrate digestion is relatively simple from an enzymatic perspective. This process is somewhat complicated by the insoluble nature of fat. Rather than letting food diffuse and break down into aqueous pieces, fat will aggregate in globules and often float as it passes through the digestive tractThink about how oil separates in water.. Therefore lipid digestion is a combination of enzymatic processing, along with the solubilization needed for absorption.
The first lipid digestive enzymes are secreted in the oral cavity. This enzyme is known as Lingual Lipase and is secreted from glands underneath the tongue. While it is present in the mouth, it is only functional in the low pH environment, such as that in the stomach. Lingual Lipase tends to cleave the sn3 fatty acids from triglyceridesRecall, phospholipids do not have a fatty acid esterified at the sn3 position, that is where the headgroup is located.. This results in a diacylglycerol and a free fatty acid. Lingual Lipase is especially effective in removing medium chain fatty acids from triglyceride molecules (Jensen et al. 1983).
Gastric Lipase also functions in the stomach. This enzyme is secreted from chief cells in the stomach, and like Lingual Lipase is most active in the low pH of the stomachThough it is able to continue functioning into the small intestine.. Gastric Lipase prefers to release fatty acids from the sn1 and sn3 positions of a lipid. For both phospholipids and triglycerides this leaves a monoacylglycerol with the fatty acid still in the sn2 position. The removal of the acyl chain make the lipid much more soluble and easier to emulsify into smaller droplets.
The process of emulsification is essential to lipid absorption. Emulsification is the breaking of lipid droplets in to smaller and smaller particles. A large globule of fat is not going to be able to easily pass through a cellular membrane, so as lipids pass through the digestive tract, both enzymatic digestion and mechanical churning to make the droplets as small as possible.
Absorption and Digestion of Lipids in the Small Intestine
Most lipid digestion and absorptionIn most cases greater than 90%. occurs within the small intestine. At this point the semi-digested, emulsified lipid droplets come into contact with bile salts, which further aid in their solubilization into very tiny lipid droplets called micelles. Micelles are generally coated with an amphipathic layer of bile salts, and contain an internal core of fatty acids, and monoacylglycerols.
Bile Salts and Their Regulation
Bile salts are cholesterol-derived compounds that are generated initially in the liverWe will discuss this in the lipid transport and synthesis lectures, but cholesterol is generally made throughout the body, with excess trafficked to the liver. Since it cannot be used for fuel, cholesterol is only released in the form of bile acid secretion. In the liver through a series of enzymatic steps, cholesterol is converted into primary bile acids such as cholic acid and chenodeoxycholic acid. This step is rate-limited by an enzyme known as 7-\(\alpha\)-hydroxylase, which in turn is transcriptionally downregulated when liver primary bile acids are high (Ramirez et al. 1994)The sensing of bile acids turns out to be an emerging area of research. The actual receptors for bile acids are a transcription factor known as FXR (farnesoid-x-receptor) and a receptor on the cellular surface known as TGR5. For 7-\(\alpha\)-hydroxylase, FXR seems to be the more important regulator (Sinal et al. 2000)..
Conjugated bile acidsA third class of bile acids, known as secondary bile acids are generated in the large intestine by bacterial modification of secreted bile acids. Since these are reabsorbed and sent back the liver, the secondary bile acids, known as deoxycholic acid and lithocholic acid can then also be conjugated in the liver. Secondary bile acids comprise of about 20-40% of the total bile acid pool. Interestingly changes in bile acids are thought to play a role in the metabolic benefits of bariatric surgery (Evers et al. 2017). are generated from primary bile acids. while still in the liver, bile acids have either a glycine or a taurineTaurine is an amino acid, derived from cysteine that is not used in proteins. amino acid group added to them. This yields a conjugated bile acid, of which there are several species. This new bile acid is very amphipathic, with a charged group from the amino acid on one end and the modified cholesterol on the other end. This makes bile acids very effective in interacting with and solubilizing dietary lipids.
Bile salts are efficiently reabsorbed, after their lipid cargo has been absorbed with up to 95% of bile salts being reabsorbed via a sodium co-transporter in the terminal illeum. One approach therefore to remove cholesterol from the blood stream is bile acid sequestrants that impair the uptake of bile salts, and thus (indirectly) the excretion of cholesterol.
Uptake of Lipids in the Small Intestine
After a brief diversion about bile salts and their role in generating lipid micelles lets return to the small intestine where we now have partially hydrolyzed triglycerides and phospholipids. A third Lipase, called Pancreatic Lipase is secreted into the small intestine, it again is specific to the sn1/sn3 positions but compared to Lingual Lipase, has stronger activity towards long chain fatty acids (Jensen et al. 1983). Pancreatic Lipase activity is dependent on a coenzyme called colipase. It is secreted as a precursor called procolipase, and is activated by trypsin-mediated cleavageThis is common theme that will come up again in the protein digestion unit wherein trypsin is also secreted as an inactive precursor and is activated by cleavage by enteropeptidase.. Recall that at this stage many lipids are solubilized within bile-salt containing micelles. The presence of colipase allows Pancreatic Lipase to be active even on lipids contained within the micelles. More details about how colipase can help Pancreatic Lipase function can be found in Van Tilbeurgh et al. (1999).
Cholesterol and Phospholipids are also digested in the small intestine. We have been focusing mainly on triglycerides, but two more enzymes in the small intestine are important for the absorption of phospholipids and cholesterol. For phospholipids, the key enzyme is Pancreatic Phospholipase. This is a class A2 phospholipaseThis means that it releases the fatty acid from the sn2 position of a phospholipid, leaving a fatty acid at the sn1 position, and the headgroup at the sn3 position.. At this stage the phospholipid is known as a lysophospholipid. For cholesterol if the cholesterol is esterifiedMeaning it has a fatty acid conjugated to it. this is removed by an enzyme termed Cholesterol Esterase. Similarly to a lysophospholipid, cholesterol is now in a more amphipathic form allowing for better absorption.
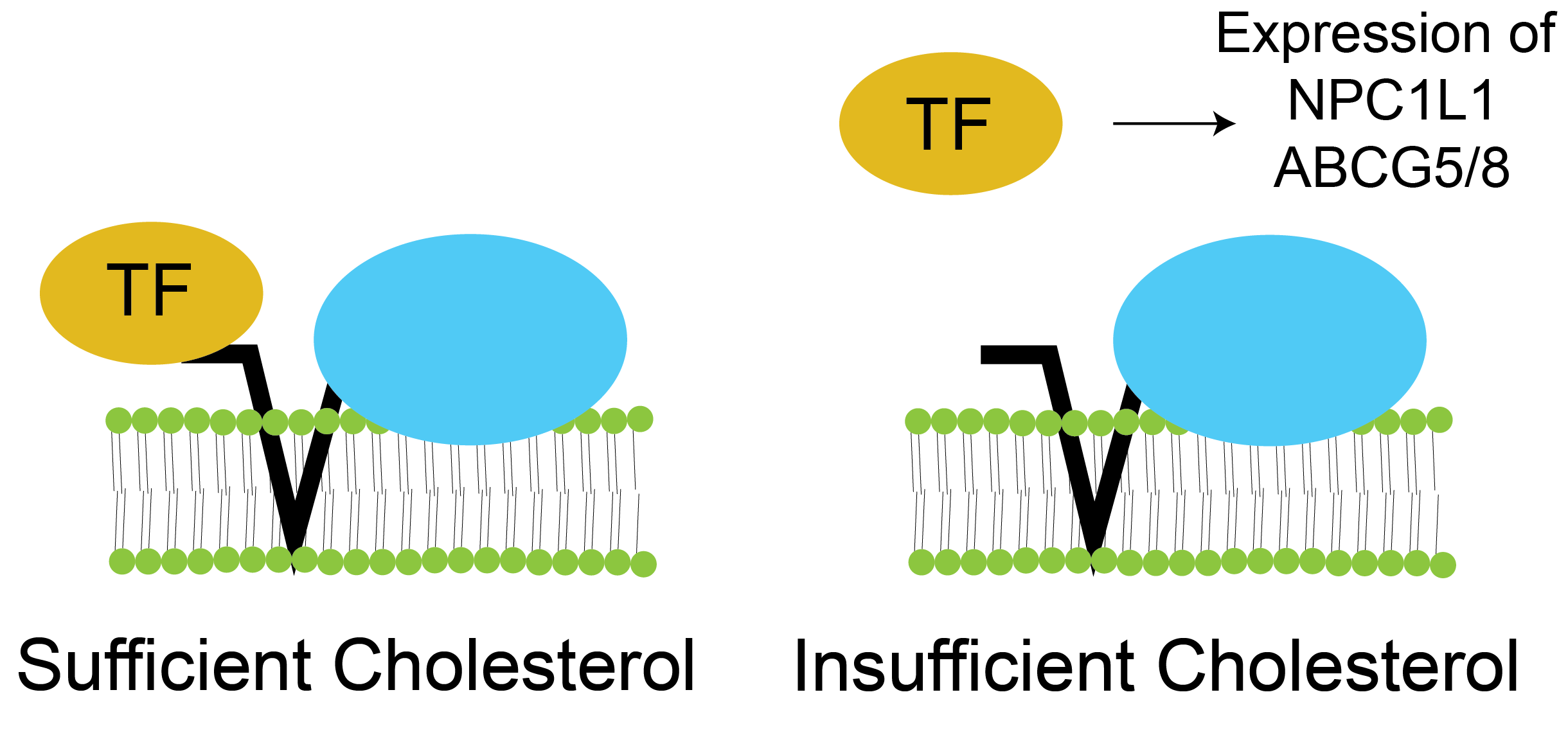
Cholesterol uptake is mediated by the NPC1L1 transporter. The predominant pathway for cholesterol uptake is via a steroid transporter called NPC1L1Niemann-Pick C1 Like 1, not the most useful name, it is based on homology with the lysosomal cholesterol transporter that causes Niemann-Pick disease, a disorder which involves lipid accumulation within lysosomes of cells.(Altmann et al. 2004; Iqbal and Hussain 2005). Plant sterols, which look very similar to cholesterol are imported along with cholesterol, but then are specifically exported back into the gut lumen via ATP-dependent transporters ABCG5/8Why do you think this step requires active transport?. Cholesterol uptake is reduced when enterocyte cholesterol levels are high. For example, bile acid synthesis deficiencySuch as mutations in 7-\(\alpha\)-hydroxylase, prevents bile acid production and secretion. results in a dramatic decrease in cholesterol absorption to balance the limited release (Repa et al. 2000; Wang 2007). The mechanism for this is via a transcription factor called SREBP2. Normally SREBP2 increases the levels of NPC1L1. When cholesterol levels are high, SREBP2 is inhibited, and cholesterol uptake is reduced (see Figure ↑. The upshot of this is that when endogenous cholesterol levels are high, dietary cholesterol absorption is reduced. This is likely one reason why dietary cholesterol intake does not strongly modulate blood cholesterol levels.
The remaining micelles contain fatty acids, monoacylglycerolsWith a fatty acid still in the sn2 position and lysophospholipids. These lipids are absorbed across the apical membrane of enterocytes. The precise mechanism seems to be a combination of passive transport and passive diffusion of micelles within the membranes, leaving the bile salts in the gut lumen. Remember that both micelles and phospholipid membranes are very amphipathic, so one theory is that the lipid containing micelles just passively pass through the membrane. Another thought is that they are bound to surface receptors then endocytosedThis means that they are actively brought into the enterocyte via a biological import process. Either way, the majority of lipids are taken out of the gut lumen into the microvilli in the small intestine.
Short Chain Fatty Acid Absorption
Both short and medium chain fatty acids are more or less soluble in water and therefore do not require micelles for transport into the enterocytes. These fatty acids are thought to be passively absorbed in the small intestine after digestion.
Short chain fatty acids are derived from fiber. On the other hand, short chain fatty acids are more rare in our diet than long chain fatty acidsThese include things like acetate, butyrate and propionate. If ingested directly these can be absorbed by enterocytes, but most SCFA’s are generated by our microbiome in the large intestine. These bacteria use undigested fiber and unabsorbed proteins and peptides as fuel. The major products of these reactions are SCFA (Cummings et al. 1987). Colon epithelial cells can take up these metabolites, and use these SCFA as a fuel source, preferring butyrate. Acetate and propionate typicaly enter the circulation. While this may seem like a minor event, SCFA’s provide about 10% of our total caloric requirements and are a major way by which the gut microbiome can affect our energy balance. For more details about SCFA and bacterial physiology refer to a recent review by Koh et al. (2016).
Endocrine Control of Lipid Digestion and Absorption
We have mentioned several pancreatic and biliary secretions that aid in digestion of lipids. The major regulators are cholecystokinin and secretin. CCK is released from the lower duodenum and acts at several places including the gallbladder and pancreas. In the gallbladder, CCK promotes contraction of the gallbladder, and relaxation of a sphincter connecting the bile duct to the duodenumThis is known as the Sphincter of Oddi.. This results in excretion of bile salts into the small intestine. As we have discussed previously CCKSecreted from enteroendocrine cells, also known as L-cells., is activated by the parasympathetic nervous system and inhibited by the sympathetic nervous system. At the same time, CCK promotes the release of sodium bicarbonate and Pancreatic Lipase and colipase from the pancreas. This process is also aided by secretin which also promotes pancreatic juice release.
Transport of Lipids out of the Enterocytes
Lipids are absborbed as free cholesterol, monoacylglycerol, lysophospholipids and free fatty acids into the enterocyte. Each of these can be quite toxic to the cell, so they are very rapidly reconverted into storage forms (esterified cholesterol and triglycerides). This re-esterification is critically important for absorption and eventual transport to other tissues.
Within enterocytes, the neutral lipids are packaged into large lipoprotein complexes called Chylomicrons. This is the first lipoprotein complex we will discuss. These particles vary in size from quite large (like chylomicrons) to very small and denseThe high density lipoproteins or HDL we be described in the lipid transport lecture. The surface of the chylomicrons contains phospholipidsMostly phosphatidylcholine (Wood et al. 1964)., free cholesterol and specific amphipathic proteins called apolipoproteins. In the case of chylomicrons, these protein are Apolipoproteins A1 and B48. The chylomicrons are secreted from the enterocytes into the lacteals of the lymphatic system. They then bypass the portal vein at first and travel to peripheral sites for rapid utilization in tissues such as adipose and muscle. Once in circulation, chylomicrons pick up Apolipoproteins CII and E. As we will describe in the lipid transport lecture these allow for the chylomicrons to recognize and activate Lipoprotein Lipase for final delivery of fatty acids and cholesterol to peripheral tissues.
Medium and short chain fatty acids on the other hand, are secreted directly into the bloodstream. These more soluble fatty acids are directly transported via the portal vein to the liver. This is one of the reasons why gallbladder and chylomicron-generating diseases can be treated with medium chain fatty acids, and why medium chain fatty acids are rapidly converted into ketones.
Reflection Questions
A patient undergoes cholecystectomy (gallbladder removal) and now receives a continuous low-level drip of unconcentrated bile into the duodenum rather than a concentrated bolus release after a meal. Evaluate how this change affects micelle formation, Pancreatic Lipase function, and fat-soluble vitamin absorption, and suggest dietary modifications that could help manage symptoms.
Ezetimibe is a drug that blocks NPC1L1-mediated cholesterol uptake in enterocytes. Using your knowledge of SREBP2 regulation and endogenous cholesterol synthesis, analyze why ezetimibe alone often has only a modest effect on circulating cholesterol, and explain the mechanistic basis for why combining ezetimibe with a statin produces a synergistic reduction.
A child with cystic fibrosis has severely impaired pancreatic secretion, including near-absent Pancreatic Lipase and colipase. Apply your knowledge of lipid digestion to predict: (a) which classes of dietary lipids would be most severely affected and why, (b) what GI symptoms and nutritional deficiencies would result, and (c) why medium chain triglycerides are used clinically to supplement fat intake in these patients.
A patient with elevated LDL cholesterol is advised to increase soluble fiber intake. Using your knowledge of bile acid enterohepatic circulation, fiber’s adsorptive properties, and SREBP2-mediated regulation of cholesterol uptake, explain the chain of events by which co-ingestion of soluble fiber with a cholesterol-containing meal reduces net cholesterol absorption — and predict why this effect requires consistent daily fiber intake rather than occasional high-fiber meals.