Glycogen Metabolism
Learning Objectives
Evaluate how the structure of glycogen allows for compact but accessible storage of glucose molecules.
Understand how glycogen synthesis and glycolysis are regulated by intracellular metabolites.
Explain how protein phosphorylation regulates Glycogen Synthase and Glycogen Phosphorylase activities and how extracellular signals affect glycogen metabolism.
Assess the tissue-specific roles of insulin, adrenaline and glucagon in glycogen storage.
Distinguish between the functions of glycogen in liver, adipose and muscle and evaluate how alterations in glycogen metabolism affect the physiological functions of these tissues.
Explain how glycogen storage diseases can occur and how specific genes result in different pathophysiologies depending on both the gene, and tissue where it is expressed.
Key Concepts and Vocabulary
Glycogenesis
Glycogenolysis
Protein Phosphorylation
Allosteric Regulation
Adrenergic Signaling
Branch Points
Inborn Errors of Metabolism
Structure and function of glycogen
Glycogen is a homopolymer of glucose units connected to each other by \(\alpha\)1-4 or \(\alpha\)1-6 glycosidic linkagesThis nomenclature refers to a link between the #1 position of one molecule of glucose (the anomeric carbon) and either the #4 or the #6 position on the next glucose.. This allows for many molecules of glucose to be compactly stored in the cell, and then made available upon energy or glucose demand. A series of \(\alpha\)1-4 bonds result in a more or less straight chain of glucose molecules, while a \(\alpha\)1-6 linkage results in a branch point. A single glucose monomer can have both an \(\alpha\)1-4 and \(\alpha\)1-6 linkage, and in glycogen these are typically spaced 8-12 glucose molecules apart (see Figure ↑). Compare this to the structure of the major dietary polysaccharides we discussed earlier in this unit (Table ↑).
| Molecule | Main Linkages | Branches |
|---|---|---|
| Glycogen | ()1-4 | ()1-6 (every 8-12) |
| Amylose | ()1-4 | none |
| Amylopectin | ()1-4 | ()1-6 (every 24-30) |
| Cellulose | ()1-4 | none |
Role of glycogen branching
The branched structure of glycogen means that a single macromolecule can be both very compact, but have many free glucose endsThese free ends are known as reducing ends.. If glycogen was totally linear then glucose could only be released one at a time from the one free end of a glycogen molecule. By having many branch points, multiple enzymes that liberate glucose from glycogenHumans only have two enzymes that can do this, one is Glycogen Phosphorylase in human tissues, the other is \(\alpha\)-amylase in our digestive tract. can release glucose from the many reducing ends at the same time. This allows for rapid mobilization of glucose when needed.
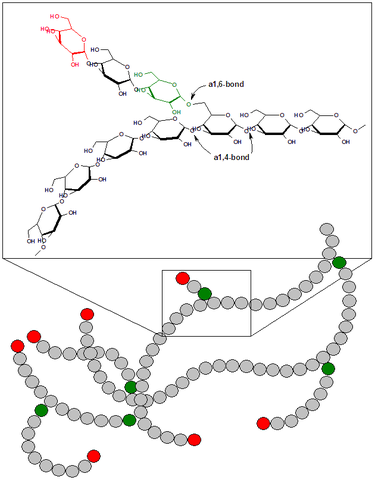
Because branch points need to be removed by a separate enzymeknown as glycogen debranching enzyme or amylo-alpha-1, 6-glucosidase (encoded by the AGL gene)., overly branched glycogen is extremely compact, but its digestion may be limited by how well Glycogen Phosphorylase and the glycogen debranching enzyme can work in concert. Defects in debranching enzyme result in a glycogen storage disease characterized by an inability to eliminate these branch points (see the section on Glycogen storage diseases below).
Different tissues use glycogen for different purposes
Glycogen is an easily accessible source of glucose for many tissues. The content of glycogen ranges from 1-3% of total weight in muscle tissue to up to 10% of total mass in a well fed liver. Since glycogen is very hydrophilic, it is estimated that approximately 3g of water are bound for every gram of glycogen (Olsson and Saltin 1970; Fernández-Elías et al. 2015). This is one reason why very low carbohydrate diets can result in rapid, initial weight loss, as glycogen is rapidly depleted. Based on what we have already learned about how glucose and phosphorylation differs between cells, different tissues use glycogen for slightly different reasons. In general, muscle cells use glycogen when energy is needed, for example during exercise. Liver cells on the other hand store glycogen to make glucose available for itself and other tissues, rather than for energy.
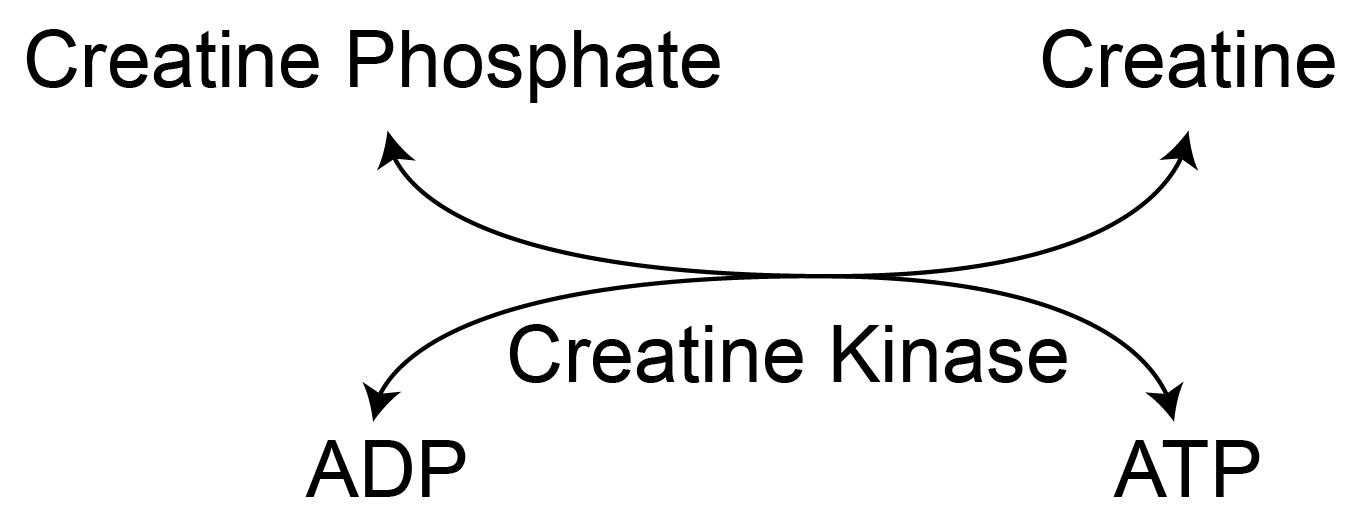
Glycogen is an important energy source. Muscle tissue can have dramatic and rapid depletions of ATP in response to exercise. The initial resource to replenish ATP is the creatine phosphate system described in Figure ↑. Think of creatine phosphate like the battery on your laptop, and can supply (or store) excess ATPWe will discuss this in the unit on non-protein compounds derived from amino acids later in the semester.. The amount of creatine phosphate can be quickly depleted, so muscle cells next turn to glycogen for glucose. These glucose molecules typically undergo glycolysis (depending on the muscle fiber type) then the TCA cycle and electron transport chain to make ATPRecall, the absence of glucose-6-phosphatase means that liberated glucose-6-phosphate will not be dephosphoryated for extracellular release.. As we will describe below, the primary signals for glycogen metabolism in muscle are energy dependent.
Glycogen plays an important role in endurance exercise. During prolonged exercise, muscle glycogen can be dramatically depleted. To prevent this, athletes often ingest carbohydrates during exercise in order to continuously provide energy to the contracting muscles. Another approach is to deplete glycogen levels prior to exercise, then eat a large carbohydrate rich meal. This results in more glycogen storage than the normal fed states, a condition known as glycogen super-compensation. These extra glycogen stores are thought to fuel a longer sustained effort during exerciseIf you are interested in exploring this more, consider the bonus group assignment Glycogen Stores and Sports Nutrition, or doing a research project on this topic.. For more information on this concept see Hawley et al. (1997).
Glycogen from the liver plays an important role in glucose homeostasis. The liver on the other hand mobilizes glycogen in order to maintain glucose levels in blood. Whether due to fasting or exercise the body needs to make glucose available for many tissues. This is especially important for the brain, which is very poor at converting fatty acids into ATP. As such, glycogen levels in the liver are generally controlled by indicators of glucose levels, such as glucose-6-phosphate. Unlike the muscle, once glycogen is catabolized into glucose-6-phosphate it can be dephosporylated into glucose and released from hepatocytes for transport to other tissues.
Regulation of glycogen synthesis
Glycogen is stored when glucose and energy are plentiful. After a typical meal, in a healthy person glycogen levels increase by about 50% peaking about 4h after a meal (Taylor et al. 1996). This is due to a combination of increased glucose availability and the postprandial actions of insulin. Glycogen is synthesized starting from Glucose-6-phosphate via the following series of reactions:
\[\begin{equation} Glucose-6-phosphate \rightleftharpoons Glucose-1-phosphate \end{equation}\]
\[\begin{equation} Glucose-1-phosphate + UTP \rightleftharpoons UDP-Glucose + PPi \end{equation}\]
\[\begin{equation} UDP-Glucose + Glycogen_n \rightarrow Glycogen_{n+1} \end{equation}\]
There are two important things to note about this process, one is that the first two reversible steps mean that the levels of G6P are extremely important in this processRecall from the lecture on glycolysis that G6P levels are controlled by the levels of glucose in the cell, the activity of hexokinase/glucokinase and the activity of PFK1, the rate limiting step in glycolysis.. The second is that by using UTP to activate glucose, this is an energy consuming process. This means that there is an energetic cost (the equivalent of one ATP phosphodiester bond) to storing glycogenPut another way, this means that to store and then release 1000 molecules of glycogen, you will use up the equivalent of 1000 ATP molecules. In the end, you have used up 1000 ATP molecules but end up where you started, with available glucose. This is another example of metabolic inefficiency, and is measured as part of the energy released during diet-induced thermogenesis.. The third reaction is catalyzed by the enzyme Glycogen SynthaseThere are two isoforms of Glycogen Synthase, one expressed in muscle and brain (GYS1) and one expressed in the liver and fat, encoded by the GYS2 gene. and that is the main point of regulatory control in glycogen synthesis.
Glycogen Synthase is activated by Glucose-6-phosphate
Both isoforms of Glycogen Synthase are allosterically activated by glucose-6-phosphate as first described in the late 1950s by Leloir et al. (1959). G6P levels are increased when glycolysis is low, but glucose levels are high. This is generally a situation where nutrient levels are high, but energy demand is low. This is a good time to store extra glucose, so this makes physiological sense.
| Kinase | Signal |
|---|---|
| PKA | Adrenaline/Glucagon |
| GSK3 | Insulin (inactivates) |
| AMPK | Energy Stress |
Extracellular control and signaling
In addition to this metabolite-level control, Glycogen Synthase is also regulated by reversible protein phosphorylation (Villar-Palasí and Larner 1960)This was one of the earliest described examples of metabolic control by protein phosphorylation.. There are several protein kinases that regulate Glycogen Synthase, and as a general rule they result in the inactivation of the enzyme. These kinases are summarized in Table ↑. Insulin activates Glycogen Synthase, and it does so by dephosphorylating these sites (see Figure ↑). Part of this mechanism is by reducing the activity of the kinases (especially GSK3 GSK3 is Glycogen Synthase Kinase that phosphorylates Glycogen Synthase and inactivates it thus decreasing the rate at which glycogen is produced. Think of why insulin may deactivate GSK3. and PKA) but insulin also functions by activating protein phosphatase activity towards Glycogen Synthase. This is accomplished via a series of proteins that specifically target a protein phosphatase on the glycogen particle. The precise mechanisms by which insulin promotes this dephosphorylation are still unclear. At the same time that Glycogen Synthase is being dephosphorylated (and activated), Glycogen PhosphorylaseThe enzyme that cleaves glucose from an existing glycogen molecule is also dephosphorylated and inactivated.
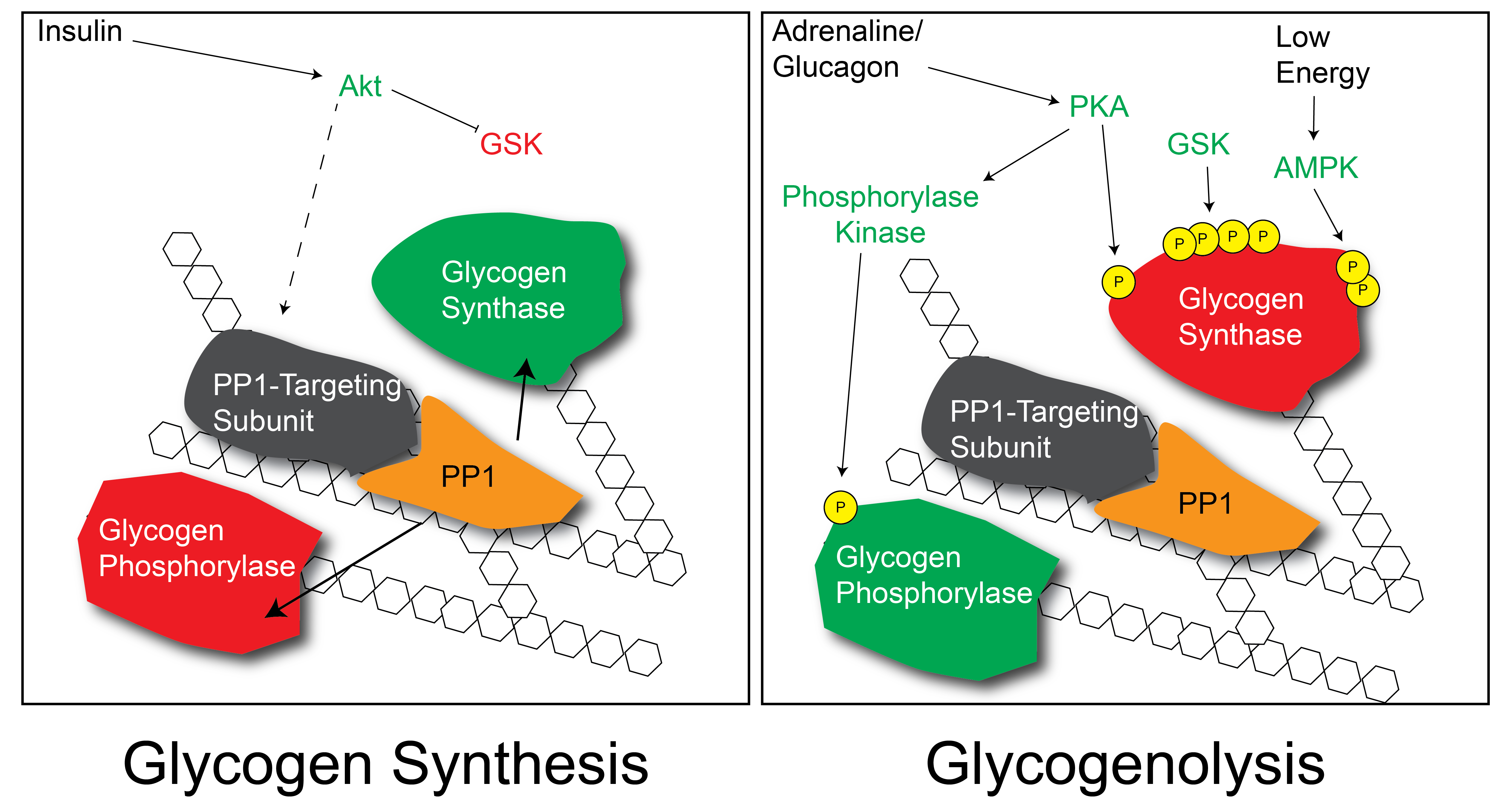
Separate from the effects of insulin on Glycogen Synthase activity, recall that insulin will also promote glucose uptake (in muscle and fat tissues). This increased glucose flux will result in more Glucose-6-phosphate in the cell and allosteric activation of Glycogen Synthase. Therefore there are at least two ways by which insulin can promote glycogenesis.
Regulation of glycogen breakdown
Glycogen is broken down to release glucose in two stepsThis mechanism is specific to the \(\alpha\)(1-4) linkages, not the branch points, which will be described later.:
\[\begin{equation} Glycogen_{n} \rightarrow Glycogen_{n-1} + Glucose-1-phosphate \end{equation}\]
\[\begin{equation} Glucose-1-phosphate \leftrightarrow Glucose-6-phosphate \end{equation}\]
The fate of Glucose-6-phosphate depends on the relative activities of PFK1, Glucose-6-phosphate dehydrogenaseLeading towards the pentose phosphate pathway. and, in the case of liver cells Glucose-6-phosphatase. Generally, in the muscle the liberated Glucose-6-phosphate enters glycolysis whereas in the liver it is dephosphorylated and released as glucose when energy is needed in the body.
Intracellular control
The first, and rate limiting step of glycogenolysis is catalyzed by an enzyme named Glycogen PhosphorylaseThere are three isoforms of this gene, PYGL, PYGM and PYGB which are expressed in the liver, muscle and brain, respectively.. Glycogen Phosphorylase is allosterically activated by AMP. This activation by AMP is blocked by the presence of ATP or Glucose-6-phosphate. AMP is increased when there is energy demand, so if there is a need for energy, Glycogen Phosphorylase gets activated. This can be over-ridden when ATP is plentiful (indicating a lack of energy stress) or Glucose-6-phosphate is elevated (indicating sufficient glucose levels). While all three isoforms respond similarly in direction, the muscle enzyme is much more sensitive to activation by AMP than the liver enzyme. This is due to structural differences in the AMP-binding pocket between the muscle and liver isoforms (Rath et al. 2000). As part of its reaction mechanism, Glycogen Phosphorylase also uses Vitamin B\(_6\)-derived Pyridoxal phosphate as a prosthetic groupVitamin B\(_6\) has many other roles, but has been shown to be effective in treating McArdle’s disease, a genetic deficiency in PYGM..
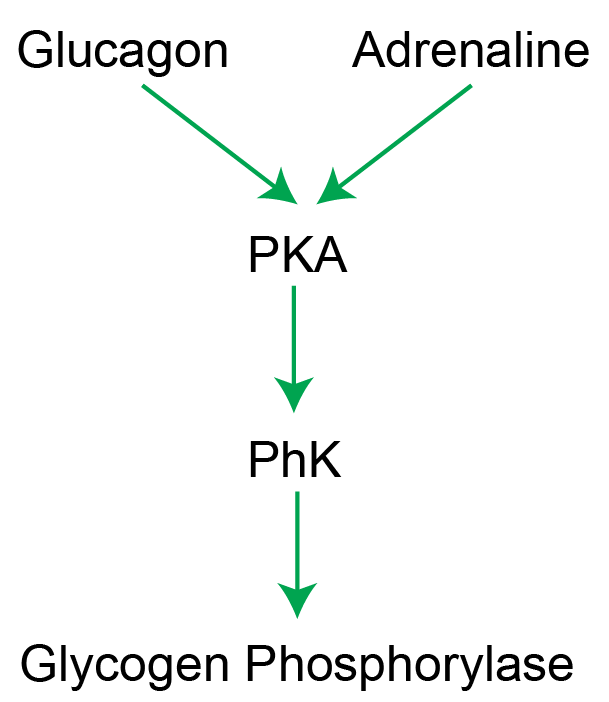
Extracellular control and signaling
The activation of Glycogen Phosphorylase by AMP can be over-ridden by protein phosphorylation by an enzyme named Phosphorylase Kinase (see Figure ↑). Once phosphorylated, the enzyme functions as if it is in the AMP-activated state. The phosphorylation of Glycogen Phosphorylase is activated by PKA dependent signaling, induced by either glucagon in the liver or adrenaline in liver, muscle and other tissues. Similarly, PKA-dependent signaling phosphorylates and inactivates Glycogen synthase. This means that adrenergic signaling turns Glycogen Phosphorylase on, and Glycogen Synthase off simultaneouslyAlso, insulin turns Glycogen Phosphorylase off, and Glycogen Synthase on.. This makes sense since adrenergic signals would want to increase release of glucose and reduce glycogen storage. A summary of the effects of reversible protein phosphorylation on the enzymes of glycogen metabolism is shown in Table ↑.
Regulation of glycogen branching and removal of branch points
The generation and removal of glycogen branch points is an important part of glycogen metabolism. Glycogen Phosphorylase can only cleave at \(\alpha\)1-4 bonds and cannot proceed past \(\alpha\)1-6 linkages. Whenever an \(\alpha\)1-6 linkage is encountered, the Glycogen Debranching Enzyme is recruited, which removes the \(\alpha\)1-6 link and allows for Glycogen Phosphorylase to proceed. Currently, there is no strong data suggesting that either Glycogen Branching or Debranching Enzymes are regulated by metabolites, or hormonal signals, but inappropriate activity can result in under- or over-branched glycogen.
| Enzyme | Effects of Phosphorylation |
|---|---|
| Glycogen Synthase | Inactivates - Less Synthesis |
| Glycogen Phosphorylase | Activates - More Breakdown |
Glycogen storage diseases
There are a variety of rare, heritable defects which result in abberant glycogen metabolism (see Table ↑). Some of these result in an inability to synthesize glycogen, while others prevent glycogenolysis, resulting in pathologically large particles of glycogen resulting in cell death. Some common glycogen storage diseases, and the affected enzymes are below.
.
| Disease | Enzyme | Predicted Glycogen Levels |
|---|---|---|
| Fanconi-Bickel syndrome | SLC4A2 | |
| von Gierke’s disease | G6PC | |
| Tarui’s disease | PFKM | |
| GSD Type 0 | GYS2 | |
| Cori’s disease | AGL | |
| Andersen disease | GBE | |
| McArdle disease | PYGM | |
| Hers’ disease | PYGL | |
| GSD type IX | PHKA1/2 |
Reflection Questions
A patient with type 1 diabetes consumes a large carbohydrate meal after a prolonged bout of exercise. Analyze what happens to glycogen synthesis in liver and muscle compared to a healthy person, tracing the effects through glucose uptake, glucose-6-phosphate levels, and Glycogen Synthase phosphorylation state. Then explain why inadequate glycogen repletion after exercise creates a risk of fasting hypoglycemia hours later — and why this delayed risk is particularly dangerous overnight.
Von Gierke’s disease is caused by loss of glucose-6-phosphatase in the liver. Evaluate why this specifically impairs blood glucose homeostasis during a fast even though hepatic glycogen can still be mobilized, and explain why the same enzyme deficiency would have a different metabolic consequence in muscle tissue.
An endurance athlete practices glycogen supercompensation before a marathon: first depleting glycogen with hard exercise, then consuming a large carbohydrate meal. Apply your knowledge of both allosteric and hormonal regulation of Glycogen Synthase to explain mechanistically why glycogen stores after this protocol can exceed normal fed-state levels.